Common features of arthropods
Type of arthropods (arthropoda)
Among all groups of invertebrate animals, the type of arthropods is distinguished by the greatest variety of adaptations to the most diverse conditions of existence, an amazing wealth of forms and a huge number of species. The number of arthropod species approaches 3 million and far exceeds the number of species of all other types of animals and plants combined, including microorganisms. Insect species are especially abundant, accounting for more than 90% of known arthropod species.
It is precisely the subtle adaptations to the use of all kinds of habitats, all kinds of food sources, often inaccessible to other animals, that explain the diversity and abundance of arthropod species.
It is difficult to find such places where there would be no arthropods. They live in all seas and oceans - both in the water column, and at the bottom, and in the soil at different depths; they also live in all kinds of fresh water bodies - not only in rivers and lakes, but also in small drying puddles, in underground waters and caves, in accumulations of water in hollows of trees. Some species have adapted to life in very salty waters, others in hot springs. It seems that all the diversity of the aquatic environment on the surface of our planet has been mastered by arthropods.
A large army of representatives of this type has also adapted to life outside the water - on land, where arthropods are especially diverse. The entire soil layer, permeated with plant roots, is also abundantly populated by arthropods - in many forest and meadow soils, a million ticks and springtails per 1 m2 are not uncommon. A huge number of arthropods live on the surface of various parts of plants and inside plant tissues.
Arthropods can be found in all climatic zones. Their number and variety are enormous in the tropics, but they also exist in the high latitudes of the Arctic and on the coast of Antarctica, where new species of ticks and springtails have recently been discovered. Arthropods can also be found in the driest deserts. Figuratively, we can say that arthropods are a type of animals that, in terms of completeness of mastering the surface of the globe, has no equal.
According to many features of the structure, arthropods are close to annelids, especially to polychaetes. The remarkable zoologist of the beginning of the last century, Georges Cuvier, who proposed the division of animals into types, combined annelids and arthropods into a single type of articulated animals (Articulata). It is significant that even today some reputable zoologists, for example, V. N. Beklemishev, advocate the restoration of the type of segmented. This shows that arthropods are in any case very close to polychaete annelids. From forms close to them, once upon a time, back in those geological epochs, in the deposits of which no fossil remains were preserved, the ancestors of modern arthropods separated.
Arthropods, like annelids, are bilaterally symmetrical segmented animals. The similarity of arthropods with rings is manifested not only in external segmentation. The central nervous system in more primitive arthropods is very similar to the scalene nervous system of polychaetes.
The specific features of arthropods are due to the fact that their body is covered with a very dense and durable cuticle, which often forms a very hard shell. The cuticle in arthropods is distinguished by the outer layer of cells, the so-called hypodermis, has a very complex structure and complex chemical composition. Previously, it was believed that in arthropods, the cuticle consists only of chitin. Now it turned out that, in addition to chitin, it includes many other compounds, mainly tanned proteins, and that in most arthropods no more than a third of the substances that make up the cuticle fall to the share of chitin. Chitin is a complex polysaccharide whose molecule contains nitrogen atoms. Probably, this is not a specific compound, but a group of closely related compounds, the formulas of which have not been precisely established. Chitin is elastic, durable and resistant to chemical attack. Often the cuticle contains a lot of calcium carbonate.
The strength and hardness of the cuticle of arthropods is so great that in those places where it reaches a large thickness, the covers become inflexible. The mobility of body segments relative to each other, which ensures movement, in arthropods is achieved by the fact that membranous areas with a very thin cuticle are preserved between hard areas (sclerites); such membranes on the folds form folds that straighten out when the body is stretched. Not all segments in arthropods are mobile relative to each other; very often there is a complete fusion of neighboring segments, especially in the anterior parts of the body.
In the body of arthropods, 3 sections can usually be distinguished - the head, on which the main sensory organs are located, which serve to orient in space (eyes, antennae, etc.) ”and food capture organs. Head segments in arthropods are fused. This is followed by the chest - that part of the body on which the main motor appendages are located - legs, and in insects, wings. The back part of the body is called the abdomen. In some arthropods (for example, in centipedes, and among fossils, in trilobites), all trunk segments are more or less homogeneous, and then only 2 sections are distinguished in the body of the animal - the head and trunk. In many arthropods (in arachnids, decapods), the head and thoracic regions merge, forming a cephalothorax. And in ticks, the boundaries between body parts are generally lost, the body is fused.
Arthropods move with the help of limbs.
Since their limbs, like the whole body, are covered with cuticles, their movement is also possible only when the legs are divided into segments. According to the structure of the limbs on which arthropods rely when moving on a solid surface or with the help of which they swim, the whole type got its name.
A certain number of pairs of limbs on the head in all arthropods turn into mouth organs - jaws that are used at least partially to capture or crush food. This is also a feature characteristic of the type of arthropods. The mouth parts are so modified that it is difficult to even recognize them as limbs, for example, the proboscis of a mosquito or a bug. The number of pairs of legs that serve for movement is not the same for different arthropods. Insects are characterized by the development of 3 pairs of motor limbs, for arachnids - 4 pairs, and in some centipedes the number of legs exceeds a hundred.
Often the trunk limbs serve not only as legs, but also as gills, suckers and other organs.
In arthropods, the motor muscles consist of separate muscles, the ends of which are attached to different thickened areas of the integument, separated from each other by a soft membrane. The movement of arthropods is provided not by body bends during contraction of the entire muscles of the body, as in worms, but by contractions of usually a few muscles associated with limbs or other motor appendages (insect wings, springtail fork, etc.) - Arthropod muscles have an exceptionally high ability to contract. All of them are striated, and the transverse striation of muscle fibers found under a microscope in arthropods is more pronounced than in vertebrates - the muscles of arthropods are stronger than ours, if, of course, we take into account the size. Since the shell of arthropods also serves as a place of attachment of motor muscles, it is also a skeleton called "outer" in contrast to the internal skeleton of vertebrates that is familiar to us.
The shell of arthropods became a skeleton only as a result of a long evolution. And at first, the thickening and hardening of the integument of the distant worm-like ancestors of arthropods that lived in the seas served, presumably, as an adaptation to protect internal organs from damage. Thickening and thickening of the cuticle makes it less permeable. Therefore, it is clear that only very small arthropods with thin integument, living in water or in very humid places, can breathe the entire surface of the body.
All larger arthropods, which also have denser integuments, necessarily develop special respiratory organs, the surface of which is covered with such a thin cuticle that oxygen can easily flow through it.
Arthropods living in water have gills. For breathing in water, it is important that the surface of the gills be as large as possible. Therefore, the gills are always thin leaf-like outgrowths or tufts of finger-like processes. Typically, gills in arthropods become limbs or parts of limbs. In water, due to its high density, even thin gill leaves do not stick together with each other. Therefore, on a small area of the body (in other places protected by dense integument) many leaf-shaped gills can be accommodated.
The circulatory system and cavity fluid serve to deliver oxygen from the gills to all organs remote from them. In arthropods, the circulatory system is represented by a pulsating, usually oblong vessel, the heart, located in the dorsal part of the body. Blood enters the heart through the side openings closed by valves, and is pushed out of it into the body cavity either directly or through a few large vessels - the circulatory system is not closed. In large marine arthropods with a thick shell - lobsters, horseshoe crabs - in the blood there is a respiratory pigment hemocyanin, similar in chemical composition to our hemoglobin, but giving the blood not red, but a bluish color: hemocyanin does not contain iron, as in hemoglobin, and copper. Hemocyanin easily attaches oxygen during the passage of blood through the gills and gives it to the internal organs. Since arthropods have an open circulatory system, it is more correct to call them “blood” “hemolymph”, but zoologists also widely use the term “blood”.
On the open land surface, neither skin respiration nor respiration with the help of gills is possible; easily permeable areas of the integument let water through and quickly dry out, losing their permeability. Therefore, aquatic organisms taken to land die from loss of water or from a lack of incoming oxygen. On land, only hidden-living small arthropods living in places where the air is always saturated with water vapor, for example, in the soil, can breathe with the entire surface of the body (small mites, springtails, symphylls). Life on land outside of wet shelters requires the development of evaporative-impervious covers, and such covers also do not allow oxygen to pass through.
Terrestrial arthropods develop special respiratory organs - either "lungs", which are deep sac-like protrusions with folded walls, opening outwards only with small holes (in spiders, scorpions), or (in most terrestrial arthropods) trachea. The tracheae are thin, usually profusely branching tubes that open outward through a small inlet ("spiracle") on the surface of the body. The branches of these tubules are washed by the abdominal fluid, and the smallest branches reach individual cells. From the thin tubes of the trachea, water evaporates very slowly through the spiracles, and its losses are insignificant, and oxygen penetrates through them quickly and easily.
Already distant ancestors of different groups of arthropods have adapted to life in different environments. It is possible to distinguish a group of aquatic arthropods, which include crustaceans, allocated to the subtype of gill-breathers. True, among modern crustaceans there are those that live on land (woodlice), but by all indications they are very close to their aquatic relatives. Another subtype - tracheal breathing - is represented by typically terrestrial arthropods - various groups of centipedes and insects. They are perfectly adapted to life on land. Their integuments contain wax and fat-like substances that protect the body from drying out, and all organ systems associated with water consumption - excretory and respiratory - act so that moisture is minimally consumed. Among insects there are individual species that spend all or part of their life in water, but the structure of such aquatic insects clearly shows their origin from land-dwelling ancestors.
Over hundreds of millions of years of history, these groups have become isolated in connection with adaptations to life in two different environments - in water and on land.
The third subtype of arthropods - chelicerae - differs very significantly from both crustaceans and tracheal-breathers. Among the chelicerae there are also aquatic representatives - horseshoe crabs that live in the seas, and terrestrial - arachnids.
In the fossil state, many remains of chelicerates have been preserved. Their study, supplementing the study of modern representatives, shows how the division of classes proceeded as a result of adaptation to life in different environments - in water and on land. The study of all groups of arthropods convinces us that their transition to life on land was associated with the ability to develop covers that protect moisture from evaporation, to preserve water in the body, without which life is impossible.
In arthropods that are well adapted to life on land, the shell is often thinner than that of those living in water, but on the other hand, a very thin layer of wax-like and fat-like substances is deposited on its surface that do not allow water vapor to pass through. This layer protects the body from drying out and prevents evaporation. Therefore, all arthropods living openly on the land surface have special respiratory organs. Those arthropods that do not have special respiratory organs, which breathe with the entire surface of the skin, are forced to live in wet shelters - in the soil, in the forest floor, etc. etc., only sometimes at night and after heavy rains showing up on the surface.
Arthropods that are best adapted to life on land most often develop a tracheal system. Not only the respiratory, but also other organ systems change in arthropods with the transition to life on land.
The removal of dissolved harmful products of protein breakdown in aquatic arthropods occurs directly into the external environment through the openings of the excretory organs, which open at the base of the limbs of the head or thoracic region. And in all arthropods that are best adapted to life on land - in spiders, centipedes, insects - excretory tubes ("Malpighian vessels") open at the beginning of the hindgut. As undigested food residues and excretory products pass through it, both from one and the other, the walls of the posterior intestine absorb water, which is so difficult for land dwellers, and excrement and metabolic products are excreted from the intestine in a dehydrated state.
Arthropods adapted to life on land have developed the ability to store a lot of fat in special formations - in the fat body. The main role of the fat body is to supply it with ... water during critical periods when the body cannot receive water from the outside!
When fats are oxidized, a lot of water is formed (the so-called "metabolic water"), which goes to cover the moisture consumption of animals. In this respect, insects are similar to camels, in which, during long passages through the desert, water is formed during the oxidation of fat reserves in the hump. Aquatic groups of arthropods (crustaceans, horseshoe crabs) do not have a fat body.
The outer shell of arthropods is so dense that it does not stretch when hardened. Therefore, the growth of arthropods is accompanied by molts. In a growing animal, the old carapace becomes tight, bursts, usually first on the back, and is shed. And only in a short period, until the new shell formed under it hardens, the animal can increase in size. During the shedding of old integuments, which come off not only from the entire outer surface of the body, but also from all limbs, including the mouth organs, as well as from the anterior and posterior intestines and from all respiratory organs, arthropods are lethargic, inactive, not feed and try to hide in any shelters.
Growth in arthropods is not a continuous process, but a stepwise one; body size increases in leaps and bounds - immediately after molting. In many arthropods, during molting, a greater or lesser change in body structure occurs. Often an adult animal is adapted to a completely different way of life than a young one, and therefore individuals of different ages (the period between molts is called the age of arthropods) are even completely different from each other. In this case, development is accompanied by metamorphosis. The transformation of a caterpillar into a chrysalis and a chrysalis into a butterfly is a well-known example.
The way of life of different arthropods is not the same. Many of them are characterized by very complex behavior, which is always associated with a high development of the sense organs and the central nervous system.
Most arthropods have well developed eyes. The eyes of arthropods are simple, having one lens, and complex, or faceted. In the higher representatives of all subtypes, the eyes are complex, faceted, consisting of many small eyes located tightly next to each other. Sometimes the number of ocelli (the so-called ommatidia) in a compound eye is measured in several thousand (Fig. 185). Compound eyes are characteristic of higher crustaceans, horseshoe crabs, and insects. Trilobites living in the Paleozoic era also had such eyes - arthropods that completely died out at the end of this era. Each ommatidium perceives only one point of the object in front of it (Fig. 186). As a result, the compound eye gives an image consisting of a large number of individual points, i.e., mosaic (like a tone cliché in a book) and straight (Fig. 7, see Vol. 3, Table 18). 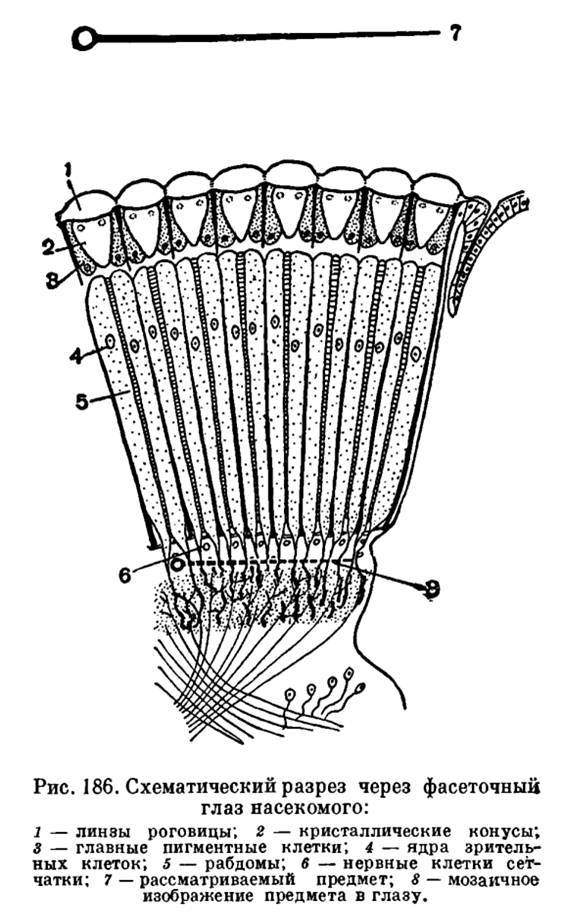
Recall that the image received in our eye is inverted and whole!
 The eyes of arthropods, it turns out, can perceive not only those rays that are visible to our eyes, but also the ultraviolet part of the spectrum. For example, many flowers that appear uniformly white to us turn out to be variegated to insects; such flowers have an ultraviolet pattern that is not visible to us (see Vol. 3, Table 19).
The eyes of arthropods, it turns out, can perceive not only those rays that are visible to our eyes, but also the ultraviolet part of the spectrum. For example, many flowers that appear uniformly white to us turn out to be variegated to insects; such flowers have an ultraviolet pattern that is not visible to us (see Vol. 3, Table 19).

Both terrestrial and aquatic arthropods are able to distinguish between ordinary light and the so-called polarized light (those light rays whose electromagnetic oscillations occur in the same plane). Since the scattered light of the sky is characterized by a certain degree of polarization of the rays in the meridional plane, arthropods in the sky, like a compass, can navigate both on clear and cloudy days. Thus, arthropods distinguish not only the frequency, but also the plane of oscillations of light rays. 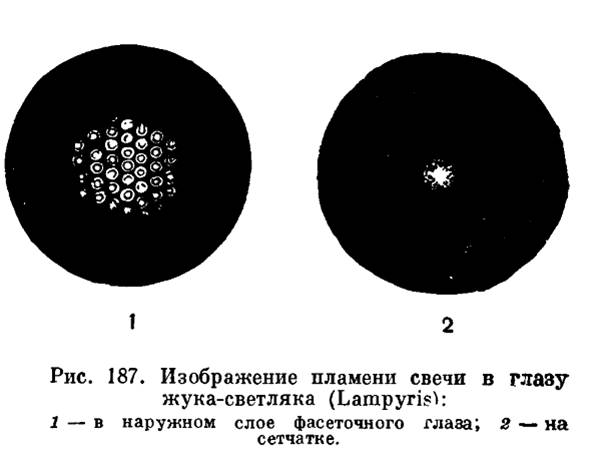
We do not feel magnetic waves. And for example, female termites in the nest lie so that the axis of their body is directed along the magnetic meridian, and if a strong magnet is placed nearby, they change their position. How arthropods perceive magnetic waves has not yet been clarified. It is noted that arthropods can also respond to short radio waves.
Hearing is also well developed in arthropods. They perceive sound waves of a certain length (usually short ones) with the help of thin auditory hairs on the surface of the body, especially on the antennae, with the help of special auditory organs, resembling a string stretched over the resonator fossa (like a string on a balalaika), and other devices. Often, terrestrial arthropods emit ultrasounds that we do not hear using various "stridulatory" organs, which, in principle, resemble a string and a bow.
Particularly remarkable is the ability of arthropods to perceive odors - negligible concentrations of many substances dissolved in water or suspended in the air. None of our chemical devices make it possible to capture such weak concentrations of a substance, and insects, for example, can navigate by them. Butterflies of scoops can fly a kilometer away to a plate with fermenting molasses, and male silkworms fly for kilometers to the smell of a box in which a female sat, which we do not feel and is close to. No less subtle is the perception of both odors and salt content by aquatic arthropods. Many terrestrial arthropods (woodlice, insect larvae) are able to detect differences in relative air humidity of the order of 1–2%.
Aquatic arthropods have well-developed balance organs.
The most universal sense inherent in all arthropods is touch. The tactile function is performed by numerous hairs on the surface of the body of arthropods. For many arthropods, especially for benthic, soil and living in the tissues of plants and animals, the desire for the largest surface of contact with solid objects is characteristic - "thigmotaxis".
The diverse "information" received from the outside world by the sense organs of arthropods requires "information processing" by the central nervous system. The brain - paired clusters of nerve cells above the esophagus - is very well developed in higher active arthropods, it has several sections associated with the main organs of orientation in space, located in the head section. The more complex the behavior of an arthropod, the more diverse the conditions that the animal encounters, and the more complex the body's responses to environmental influences, the more complex the structure of the brain. The brain in arthropods is connected by a peripharyngeal nerve ring with the ventral nerve cord. The starting point in arthropods, apparently, is the correspondence to each segment of the body of a pair of ganglia of the ventral nerve chain. But when individual segments and even parts of the body merge with each other, a decrease in the number of nerve nodes occurs, partially merging with each other, partially disappearing. The degree of dissection or fusion of the ventral chain is related to the degree of dismemberment or fusion of the body, and the concentration of nodes in the ventral nerve chain cannot at all serve as an indicator of the complexity of behavior.
The behavior of arthropods, especially during the breeding season (mating, caring for offspring), can be very complex, as can be seen from acquaintance with the descriptions of the lifestyle of different representatives of this type given below. Very often, behavior is determined by a direct response to a stimulus, such as approaching a light source ("positive phototaxis") or moving away from the light ("negative phototaxis"). Even more common are reactions to irritation by increasing or decreasing mobility (“kinesis”). For example, many insects move more actively in strong or low light and are less active in medium light. Thus, after a non-directional fast movement, they finally fall into favorable lighting conditions for themselves and stop moving.
In addition to direct standard responses to stimuli (unconditioned reflexes), arthropods are also characterized by complex chains of hereditarily fixed reflex actions (instincts). Instincts represent such complex behavior that is expedient under normal conditions for a given species, but it remains unchanged when conditions change and therefore may become inappropriate.
For example, burrowing wasps dig out a mink, bring a paralyzed insect into it, which the larva should feed on, lay an egg on the prey and close up the mink. All actions of the wasp are "expedient". But if, in the course of the activity of the wasp, when she begins to close up the mink, she takes out both the prey and the egg, the wasp completes its already unnecessary work of walling up the mink.
Along with complex forms of instinctive behavior in arthropods (higher crustaceans, spiders, higher insects), individual skills can also be traced; they can also develop conditioned reflexes. Moreover, for example, bees can transfer acquired skills to each other, exchange information. With certain body movements, the worker bee informs other bees in the hive in which direction to fly for a bribe.
Arthropods, of course, are characterized by many manifestations of higher nervous activity, which until recently was considered characteristic only of vertebrates.
The types of fertilization in arthropods are varied.
In aquatic arthropods, for example, horseshoe crabs, there is a real external fertilization, when the female lays her eggs in the water, and the male brings the seminal fluid there. Penetration of the sperm into the egg occurs during external fertilization in the external environment - in water.
Often, during external fertilization in aquatic arthropods, the male attaches "packages" with seminal fluid (spermatophores) to the excretory genital openings of females - the eggs laid are fertilized in the external environment, but the probability of fertilization of each egg in this case increases.
With the transition to life on land, the lower arthropods living in a humid environment - in the soil, in rotten wood, etc. (many mites, scorpions, false scorpions, centipedes, springtails, etc.), developed a new method of fertilization. Males deposit spermatophores into the external environment, as in external fertilization (on the ground, on the remains of plants), and females pick them up with their genital openings, and thus, the penetration of spermatozoa into the egg is carried out inside the female's body, as in internal fertilization. Such fertilization is called external-internal.
In arthropods living on the land surface in an open atmosphere, fertilization is internal: males inject seminal fluid directly into the genital openings of females. The penetration of the spermatozoon into the egg occurs in conditions protected from drying out inside the mother's body.
Certain groups of aquatic arthropods, such as barnacles, have also developed the capacity for internal fertilization, which increases fertility.
In most arthropods, oviposition is observed, live birth is rare, for example, in some Diptera. Oviparous is more often observed: the female gives birth to live young, however, the development of the young occurs due to the nutrient reserves of the egg, but not in the external environment, but in the oviducts of females (for example, in aphids).
As mentioned, the development of many arthropods occurs with a complex transformation. In those cases where the adult and the form that emerged from the egg differ sharply in structure, the young individuals are called larvae.
In sedentary or immobile marine crustaceans, mobile larvae settle. In adult winged insects, which are very mobile, the larvae are mainly the feeding stage.
In cases where eggs are rich in yolk, young arthropods emerge from them, already similar to adults. This development is characteristic of many freshwater crustaceans, arachnids and some insects.
Arthropods are ancient animals.
The most reliable data, on the basis of which it is possible to judge the origin of certain groups of animals, are provided by studies of fossilized remains of organisms or their imprints in compacted and petrified layers of silt, sand or clay, under the sediments of which the animal corpses were buried. But not from all past eras, when life already existed on Earth, fossil remains of organisms have been preserved. The deposits of the most ancient periods of the development of life on Earth were subjected to such strong changes during subsequent epochs that it is impossible to determine exactly which organisms lived on Earth about a billion years ago, any surviving animal remains from deposits older than the Paleozoic have not survived . And already in the deepest ("lower") sedimentary rocks of the Cambrian period of the Paleozoic era (deposited more than half a billion years ago), trilobites are found (see v. 3, Fig. 3) - animals with pronounced signs of arthropods, which had a developed external skeleton, dissected limbs, compound eyes and other signs of organization characteristic of representatives of the Arthropoda type. Trilobites are a class of arthropods that completely became extinct at the end of the Paleozoic era (which lasted 300-350 million years). 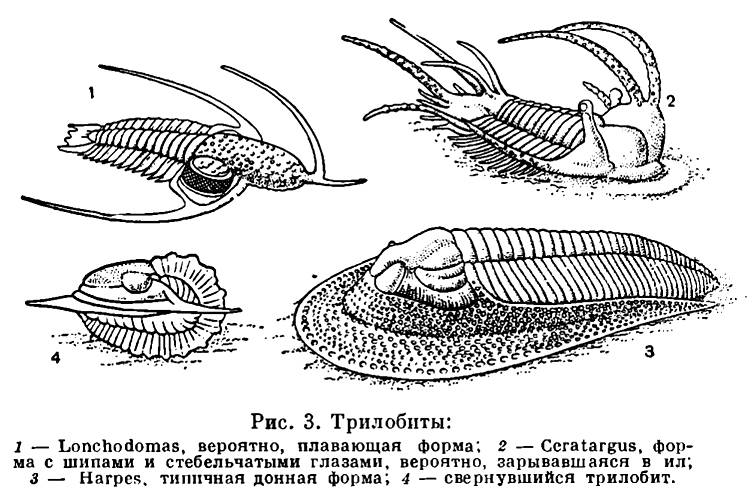
Crustaceans are also known from Cambrian deposits, some of which belong to modern orders, relatives of horseshoe crabs are known from the Silurian period (see vol. 3, table 1) and even scorpions (see vol. 3, tables 3, 1, 2) , the remains of springtails were found in the Devonian deposits, and recently the Moscow paleontologist B. B. Rodendorf established that winged insects also lived in the Devonian. 
All these data indicate that the type of arthropods separated from some animals similar to ringworms a very long time ago. Therefore, it is very difficult to accurately establish the relationship of various arthropods. There is no doubt that one of the first groups of arthropods to take shape were trilobites. In the Paleozoic, trilobites flourished, they literally swarmed in the coastal zone of the seas and oceans 500 million years ago. Trilobites have been preserved in the fossil state so well that, using some remains, it was possible not only to study their internal structure, but even to study the microscopic structure of the tissues of these animals on thin sections, and also to trace their development. And using some methods of modern chemistry, it was even possible to find out that there was hemocyanin in their blood, like in modern higher crustaceans.
Trilobites, usually classified as a separate class of arthropods, deserve a separate subtype of Trilobita.
Of the modern groups of arthropods, horseshoe crabs are apparently closest to trilobites (see vol. 3, table 1), the larvae of which are somewhat similar to trilobites, and the embryos are similar to trilobite larvae (“protaspis”—see vol. 3, Fig. 2). 
Horseshoe crabs and arachnids have a number of common structural features that allow them to be combined into a single subtype of chelicerae. (Chelicerata).
According to the nature of the respiratory organs and some other features, crustaceans (class Crustacea) also have some features in common with trilobites. However, no direct relationship between trilobites and crustaceans can be established. Crustaceans represent a special subphylum of branchials (Branchiata) in the phylum Arthropoda. And since among modern crustaceans many have a more primitive structure than chelicerae, it is customary to begin consideration of the type of arthropods with them.
The last subtype - tracheal-breathing (Tracheata) - combines various centipedes and insects, i.e., typically terrestrial arthropods.
At present, there is no doubt that insects originated from some ancient arthropods close to centipedes, and centipedes from primitive worm-like arthropods.
Probably, aquatic chelicerae descended from common ancestors with trilobites - this was another branch of the evolutionary development of arthropods. And crustaceans that have adapted to life in the aquatic environment, retaining many features of ancient arthropods, represent the third main branch of arthropod development (Fig. 188). 
Acquaintance with the type and with its diverse representatives is most convenient to begin with the subtype of the branchial, from the class of crustaceans.

Animal life: in 6 volumes. — M.: Enlightenment. Edited by professors N.A. Gladkov, A.V. Mikheev. 1970.
