Биогенетический закон: история, суть, применение. Что значит "биогенетический закон"
обобщение, согласно которому индивидуальное развитие особи (онтогенез) является как бы кратким повторением (рекапитуляцией) важнейших этапов эволюции (филогенеза) группы, к которой эта особь относится. Установлен Ф. Мюллером (1864) и сформулирован Э. Геккелем (1866).
Биогенетический закон
закономерность в живой природе, сформулированная немецким учёным Э. Геккелем (1866) и состоящая в том, что индивидуальное развитие особи (онтогенез) является коротким и быстрым повторением (рекапитуляцией) важнейших этапов эволюции вида (филогенеза). Факты, свидетельствующие о рекапитуляции (например, закладка у зародышей наземных позвоночных жаберных щелей), были известны ещё до появления эволюционного учения Ч. Дарвина. Однако лишь Дарвин дал (1859) этим фактам последовательное естественно-историческое объяснение, установив, что стадии развития зародышей воспроизводят древние предковые формы. Он рассматривал рекапитуляцию как фундаментальную закономерность эволюции органического мира. Теория естественного отбора позволила Дарвину объяснить противоречивое сочетание целесообразности строения организмов с рекапитуляцией признаков далёких предков. Немецкий эмбриолог Ф. Мюллер в 1864 подкрепил принцип рекапитуляции данными из истории развития ракообразных. Двумя годами позже Геккель придал принципу рекапитуляции форму Б. з., схематизировав при этом дарвиновские представления. Б. з. сыграл важную роль в биологии, стимулировал эволюционные исследования в эмбриологии, сравнительной анатомии и палеонтологии.
Вокруг Б. з. развернулась продолжительная и острая дискуссия. Противники Б. з. пытались истолковать Б. з. в духе механицизма, витализма или безоговорочно его отвергали. Отстаивая Б. з., дарвинисты стремились углубить его содержание и освободить от схематичности. Они критиковали представления Геккеля, ошибочно разделявшего явления эмбрионального развития на 2 неравноценные группы: палингенезы , отражающие историю вида, и ценогенезы, возникшие в качестве приспособления зародышей к условиям среды и затемняющие, «фальсифицирующие», палингенезы. Несостоятельным оказалось и первоначальное представление Геккеля о прямом порядке воспроизведения в развитии особи этапов истории вида. Было показано (в т. ч. и самим Геккелем), что гетерохронии, гетеротопии, эмбриональные приспособления, редукция и другие процессы глубоко изменяют течение онтогенеза, исключая возможность прямой рекапитуляции признаков предков. Новое освещение Б. з. получил в теории филэмбриогенеза русского биолога А. Н. Северцова. Явление рекапитуляции Северцов рассматривает под углом зрения закономерностей эволюции онтогенеза. Б. з. расценивается им как следствие эволюции, осуществляющейся путём надставки (анаболии) конечных стадий онтогенеза; ценогенезы же являются закономерным путём эволюции вида и имеют палингенетическую природу. Вопреки мнению, будто Б. з. неприложим к растениям, ряд ботаников приводил примеры рекапитуляции у растений. Обстоятельный анализ Б. з. с ботанической точки зрения был проведён советским учёным Б. М. Козо-Полянским (1937); им предложена формулировка закона рекапитуляции с учётом своеобразия онтогенеза и индивидуальности растений. Дальнейший прогресс представлений о рекапитуляции, подтвердивший ограниченность геккелевской трактовки Б. з., связан с успехами эволюционной морфологии, экспериментальной эмбриологии и генетики, которые обобщены в учении И. И. Шмальгаузена об организме как целом в индивидуальном и историческом развитии.
Лит.: Дарвин Ч., Происхождение видов..., Соч., т. 3, М., 1939; Мюллер Ф. и Геккель Э., Основной биогенетический закон, М.≈ Л., 1940; Козо-Полянский Б. М., Основной биогенетический закон с ботанической точки зрения, Воронеж, 1937; Северцов А. Н., Морфологические закономерности эволюции, М.≈Л., 1939; Шмальгаузен И. И., Организм как целое в индивидуальном и историческом развитии, М.≈Л., 1942; Мирзоян Э. Н., Индивидуальное развитие и эволюция, М., 1963.
Э. Н. Мирзоян.
Б. з. в психологии. В связи с внедрением в психологию идей эволюционной биологии в конце 19 ≈ начале 20 вв. были предприняты попытки использовать Б. з. для объяснения механизма смены стадий в развитии психических функций и форм поведения у животных и человека. Психологи, стоявшие на этой точке зрения ≈ С. Холл, Дж. Болдуин (США), П. П. Блонский (СССР) и др., ≈ утверждали, что имеется соответствие между эволюцией всего живого, в частности историческим развитием общества, и индивидуальным развитием ребёнка (например, соответствие между поведением первобытного человека и дошкольника, поведением человека античной эпохи и младшего школьника и т.д.). Такой подход явился одной из основ педологии. Научный анализ развития детей вскрыл ошибочность этой гипотезы и показал, что ребёнок усваивает опыт человечества не на основе Б. з., а под воздействием обучения и воспитания.
М. Г. Ярошевский.
Википедия
Биогенетический закон
по Геккелю. Рисунок из книги Ремане (1892), воспроизводящий исходную иллюстрацию Геккеля
Биогенетический закон сыграл заметную роль в истории развития науки, однако в настоящее время не признается современной биологической наукой.
БИОГЕНЕТИЧЕСКИЙ ЗАКОН (греческий bios жизнь, genetikos относящийся к рождению, происхождению) - комплекс теоретических обобщений, описывающих связь между индивидуальным и историческим развитием живых организмов.
Б. з. был сформулирован в 1866 г. нем. зоологом Э. Геккелем (Е.Н. Haeckel): «Ряд форм, через которые проходит индивидуальный организм при своем развитии, начиная от яйцеклетки и кончая вполне развитым состоянием, является кратким, сжатым повторением длинного ряда форм, пройденного животными предками того же организма или родовыми формами его вида, начиная с древнейших времен так наз. органического творения, вплоть до настоящего времени», т. е. «онтогенез есть быстрое и краткое повторение филогенеза».
Основой для создания Б. з. послужила работа Ф. Мюллера «За Дарвина» (1864), в к-рой было показано, что филогенетически новые признаки взрослых организмов возникают в результате изменения онтогенеза у потомков - удлинения или отклонения от онтогенеза предков. В обоих случаях меняется строение взрослого организма.
По мнению Геккеля, филогенез происходит путем суммирования изменений взрослого организма и сдвига их на более ранние стадии онтогенеза, т. е. филогенез является базой для онтогенеза, к-рому отводится роль сокращенной и искаженной записи эволюционных преобразований взрослых организмов (см. Онтогенез , Филогенез). С этих позиций все признаки развивающегося организма Геккель разделил на две категории: палингенезы (см.) - признаки или стадии индивидуального развития, повторяющие, или рекапитулирующие в онтогенезе потомков этапы филогенеза взрослых предков, и ценогенезы - любые признаки, нарушающие рекапитуляцию. Причиной ценогенезов Геккель считал вторичные приспособления организмов к тем условиям, в которых протекает их онтогенез. Поэтому к ценогенезам были отнесены временные (провизорные) приспособления, обеспечивающие выживание особи на тех или иных стадиях индивидуального развития и отсутствующие у взрослого организма, напр, зародышевые оболочки плода (собственно ценогенезы), а также и изменения закладки органов во времени (гетерохронии) или месте (гетеротопии) и вторичные изменения пути онтогенеза данного органа. Все эти преобразования нарушают палингенез и тем самым затрудняют использование данных эмбриологии для реконструкции филогенеза, ради чего, как показал А. Н. Северцов (1939), Геккелем был сформулирован Б. з.
В начале 20 в. рядом авторов было доказано, что Мюллер (F. Muller), постулировавший возникновение филогенетических изменений в результате преобразований процессов онтогенеза, правильнее, чем Геккель, объяснил соотношение между индивидуальным и историческим развитием, обоснованное в наст, время с позиций генетики. Т. к. эволюция происходит в ряде поколений, в ней имеют значение только генеративные мутации, меняющие наследственный аппарат гамет или зиготы. Лишь эти мутации передаются следующему поколению, в к-ром они меняют ход онтогенеза, благодаря чему и проявляются в фенотипе потомков. Если в следующем поколении онтогенез протекает так же, как в предыдущем, то взрослые организмы обоих поколений будут одинаковы.
На основе представления о первичности онтогенетических изменений А. Н. Северцовым была разработана теория филэмбриогенеза - описание способов (модусов) эволюционных изменений хода онтогенеза, которые приводят к преобразованию органов потомков. Наиболее распространенным способом прогрессивной эволюции органов является анаболия, или надставка конечных стадий развития. В этом случае к той стадии, на к-рой оканчивалось развитие органа у предков, добавляется новая (удлинение онтогенеза), а конечная стадия онтогенеза предков оказывается как бы сдвинутой к началу развития:
Анаболии E, F, G, H ведут к дальнейшему развитию органа и обусловливают рекапитуляцию предковых состояний (e, f, g). Следовательно, именно при эволюции путем анаболии возникает палингенетический путь онтогенеза, однако в данном случае происходит не смещение стадий онтогенеза, а дальнейшее филогенетическое развитие органа, уже существовавшего у предков.
Второй модус филэмбриогенеза - девиация, или отклонение на промежуточных стадиях развития. В этом случае развитие органа потомков начинается так же, как у предков, но затем оно меняет направление, хотя дополнительные стадии и не возникают:

Девиации перестраивают онтогенез, начиная с промежуточных стадий (с1, d2, d3), что ведет к изменению дефинитивного строения органа (E1, E2, Е3). Рекапитуляция в онтогенезе abc1d1E1 прослеживается на стадиях ab, а в онтогенезе abc1d3E3 - на стадиях abc1. Третий, наиболее редкий, модус прогрессивной эволюции - архаллаксис, или изменение первичных зачатков органов:

Архаллаксис характеризуется преобразованием самых ранних этапов онтогенеза, начиная с его закладки (а1, а2, а3), что может привести к возникновению новых, отсутствовавших у предков органов (Е1, Е2, Е3) - первичный архаллаксис, или к коренной перестройке онтогенеза органа без существенных изменений его дефинитивной структуры - вторичный архаллаксис. При этом способе эволюции рекапитуляция отсутствует.
Посредством филэмбриогенезов происходит также эволюционная редукция органов. Различают два типа редукции: рудиментацию (недоразвитие) и афанизию (бесследное исчезновение). При рудиментации орган, нормально развитый и функционировавший у предков, утрачивает у потомков свое функциональное значение. В этом случае, по мнению А. Н. Северцова, редукция осуществляется путем отрицательного архаллаксиса: закладка у потомков меньше и слабее, чем у предков, развивается медленнее и не доходит до предковой дефинитивной стадии. В результате орган потомков оказывается недоразвитым. При афанизии редуцирующийся орган не только теряет свое функциональное значение, но и становится вредным для организма. Онтогенез такого органа, как правило, начинается и нек-рое время протекает так же, как у предков, но затем возникает отрицательная анаболия - орган рассасывается, причем процесс идет в порядке, обратном развитию, вплоть до исчезновения самой закладки.
Теория филэмбриогенеза близка к представлениям Мюллера. Однако А. Н. Северцовым был выделен модус архаллаксиса, который можно наблюдать лишь при эволюционных преобразованиях частей, а не целого организма, изучавшегося Мюллером. Советские биологи доказали, что посредством филэмбриогенезов эволюционируют не только органы, но и ткани, и клетки многоклеточных организмов. Имеются доказательства эволюции путем филэмбриогенезов не только развитых органов, но и провизорных приспособлений (ценогенезов). Выяснено также, что в ряде случаев гетерохронии имеют значение филэмбриогенезов.
Т. о., филэмбриогенез является универсальным механизмом филогенетических преобразований строения организмов на всех уровнях (от клетки до организма) и этапах онтонегеза. В то же время филэмбриогенезы нельзя считать первичными и элементарными эволюционными изменениями. В основе эволюции лежит, как известно, мутационная изменчивость. Как филэмбриогенезы, так и генеративные мутации наследуются и проявляются в ходе онтогенеза. Однако мутационная изменчивость, в отличие от филэмбриогенезов, индивидуальна (каждая новая мутация свойственна лишь той особи, у к-рой она возникла), и впервые появляющиеся мутационные изменения не носят приспособительного характера. Филэмбриогенезы, по всей вероятности, представляют собой комплексы мутаций, прошедших естественный отбор и ставших генотипической нормой. В таком случае филэмбриогенезы - вторичные преобразования, возникающие как результат сохранения и накопления мутаций, меняющих морфогенез (см.), а тем самым и отроение взрослых организмов в соответствии с изменениями окружающей среды. Естественный отбор чаще сохраняет изменения, лишь надстраивающие онтогенез, реже - меняющие промежуточные стадии и еще реже - преобразующие морфогенез с самых первых его этапов. Этим объясняется различная частота возникновения анаболий, девиаций и архаллаксисов. Следовательно, филэмбриогенез, являясь механизмом становления филогенетически новых признаков, в то же время представляет собой результат мутационной перестройки индивидуального развития.
Представления Геккеля о преимущественности филогенетических изменений над онтогенетическими и Мюллера о первичности перестройки хода онтогенеза, приводящей к филогенетическим преобразованиям строения организмов, являются односторонними и не отражают всей сложности эволюционных взаимоотношений онтогенеза и филогенеза. С современных позиций связь между индивидуальным и историческим развитием организма выражается следующим образом: «филогенез есть исторический ряд известных онтогенезов» (И. И. Шмальгаузен, 1969), где каждый последующий онтогенез отличается от предыдущего.
Библиография: Лебедин С. Н. Соотношение онто- и филогенеза, библиография вопроса, Изв. Науч. ин-та им. Лесгафта, т. 20, № 1, с. 103, 1936; Мюллер Ф. и Геккель Э. Основной биогенетический закон, пер. с нем., М.- Л., 1940; Север цов А. Н. Морфологические закономерности эволюции, с. 453, М.-Л., 1939; Северцов А. С. К вопросу об эволюции онтогенеза, Журн. общ. биол., т. 31, № 2, с. 222, 1970; Ш м а л ь г а у-зен И. И. Проблемы дарвинизма, с. 318, Л., 1969.
А. С. Северцов.
Биогенетический закон Геккеля-Мюллера описывает наблюдаемое в живой природе соотношение - онтогенез, то есть персональное развитие каждого живого организма, в определенной степени оно повторяет филогенез - историческое развитие всей группы особей, к которой он относится. Закон сформулировали, как понятно из названия, и Ф. Мюллер в 60-е годы XIX века независимо друг от друга, и установить первооткрывателя теории сейчас почти невозможно.
Очевидно, что биогенетический закон не был сформулирован сразу. Работе Мюллера и Геккеля предшествовало создание теоретической базы для закона в виде уже обнаруженных явлений и других установленных закономерностей природы. В 1828 году К. Бэр сформулировал так называемый закон зародышевого сходства. Суть его заключается в том, что эмбрионы особей, относящихся к одному биологическому типу, имеют множество схожих элементов анатомического строения. У человека, например, на определенной стадии развития у зародыша имеются жаберные щели и хвост. Характерные отличительные черты в морфологии видов возникают только в ходе дальнейшего онтогенеза. Закон зародышевого сходства во многом определил биогенетический закон: раз эмбрионы различных организмов повторяют стадии развития других особей, они повторяют стадии развития всего типа вообще.
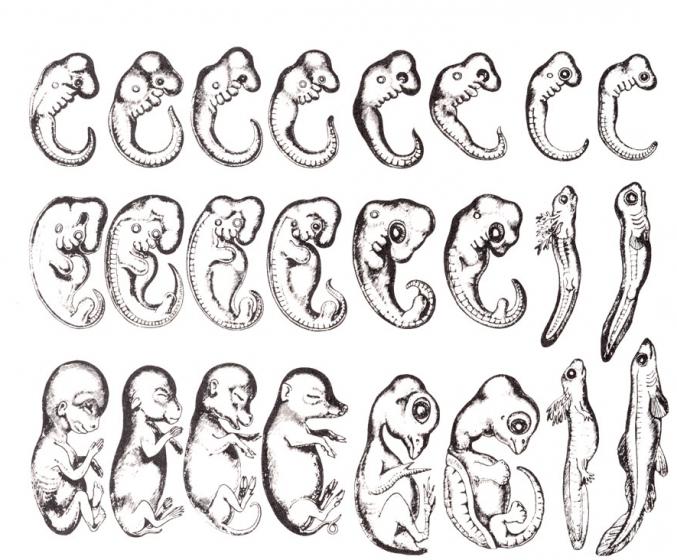
А.Н. Северцовым позднее были внесены определенные поправки к закону Геккеля-Мюллера. Ученый отметил, что во время эмбриогенеза, то есть стадии наблюдается сходство между органами именно эмбрионов, а не взрослых особей. Так, жаберные щели у схожи с жаберными щелями эмбрионов рыб, но никак не со сформировавшимися жабрами взрослых рыб.
Важно отметить, что одним из наиболее значимых доказательств дарвиновской теории эволюции считается непосредственно биогенетический закон. Формулировка его сама по себе намекает на собственную логическую связь с учением Дарвина. Эмбрион в ходе своего развития проходит через множество различных стадий, каждая из которых напоминает определенные ступени развития природы, отмечаемые с эволюционной точки зрения. Таким образом, каждая все более сложно организованная особь отражает в своем онтогенезе развитие всей живой природы с точки зрения эволюции.
В психологии также существует свой биогенетический закон, сформулированный независимо от биологического. По сути, в психологии выделяется не оформленный закон, а высказанная И. Гербартом и Т. Циллером идея о схожести развития психики ребенка с таковой у человечества вообще. Различные ученые пытались обосновать эту теорию с разных точек зрения. Г. Холл, например, прибегал непосредственно к закону Геккеля-Мюллера. Он говорил о том, что развитие ребенка, в том числе в психологическом плане, задается исключительно биологическими предпосылками и повторяет эволюционное развитие вообще. Так или иначе, на сегодняшний день идея не является однозначно доказанной. В психологии до сих пор не существует биогенетического закона как такового.
3. Факты, противоречащие биогенетическому закону
4. Связь биогенетического закона с дарвинизмом
5. Научная критика биогенетического закона и дальнейшее развитие учения о связи онтогенеза и филогенеза
6. Критика со стороны креационистов
Биогенетический закон Геккеля-Мюллера: каждое живое существо в своем индивидуальном развитии повторяет в известной степени формы, пройденного его предками или его видом.
Зародыши по Геккелю. Рисунок из книги Ремане, воспроизводящий исходную иллюстрацию Геккеля
Сыграл важную роль в истории развития науки, однако в настоящее время в своем исходном виде не признается современной биологической наукой. По современной трактовке биогенетического закона, предложенной русским биологом А.Н. Северцовым в начале 20 века, в онтогенезе происходит повторение признаков не взрослых особей предков, а их зародышей.
История создания
Фактически «биогенетический закон» был создан ещё задолго до возникновения дарвинизма.
Немецкий анатом и эмбриолог Мартин Ратке в 1825 г. описал жаберные щели и дуги у эмбрионов млекопитающих и птиц один из наиболее ярких примеров рекапитуляции.
В 1824-1826 годах Этьен Серра сформулировал «закон параллелизма Меккеля-Серра»: каждый организм в своем эмбриональном развитии повторяет взрослые формы более примитивных животных.
В 1828 году Карл Максимович Бэр, основываясь на данных Ратке и на результатах собственных исследований развития позвоночных, сформулировал закон зародышевого сходства: «Эмбрионы последовательно переходят в своем развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи». Бэр не придавал этому «закону» эволюционного смысла, однако позднее этот закон стал рассматриваться как «эмбриологическое доказательство эволюции» и свидетельство происхождения животных одного типа от общего предка.
«Биогенетический закон» как следствие эволюционного развития организмов впервые был сформулирован английским естествоиспытателем Чарльзом Дарвином в его книге «Происхождение видов» в 1859 г: «Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше более или менее затененный образ общего прародителя, во взрослом или личиностном его состоянии, всех членов одного и того же большого класса»
За 2 года до формулировки Эрнстом Геккелем биогенетического закона сходную формулировку предложил на основе своих исследований развития ракообразных работавший в Бразилии немецкий зоолог Фриц Мюллер. В своей книге «За Дарвина», изданной в 1864 году, он выделяет курсивом мысль: «историческое развитие вида будет отражаться в истории его индивидуального развития».
Краткая афористичная формулировка этого закона была дана немецким естествоиспытателем Эрнстом Геккелем в 1866 г. Краткая формулировка закона звучит следующим образом: Онтогенез есть рекапитуляция филогенеза.
Биогенетический закон Геккеля – Мюллера или основной биогенетический закон гласит: каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет в известной степени формы, пройденные его предками или его видом (филогенез).
Этот закон сыграл важную роль в истории развития науки, однако в настоящее время в своем исходном виде не признается современной биологической наукой.
По современной трактовке биогенетического закона, предложенного русским биологом А. Н. Северцовым в начале ХХ века, в онтогенезе происходит повторение признаков не взрослых особей, а их предков.
Часто с биогенетическим законом сопоставляют закон зародышевого сходства, сформулированный К. М. Бэром в 1828 году, из которого следует, что эмбрионы последовательно проходят в своем развитии от общих признаков типа ко все более специальным признакам; позднее всего развиваются признаки, указывающее на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи.
Ряд исследователей (Северцев, 1939; Шмальгаузен, 1969, Иванова – Казас, 1939) показали, что закон зародышевого сходства и биогенетический закон обуславливаются различными механизмами, и, соответственно, это два разных закона.
Северцев (1939) утверждал, что у одноклеточных онтогенез отсутствует, и только для вольвоксов признавал его наличие в самой примитивной форме.
Вслед за Северцовым большинство эмбриологов отрицает наличие индивидуального развития на клеточном уровне организации, однако, при таком подходе непонятно на базе чего сформовались морфогенетические механизмы первых многоклеточных животных?
Словосочетание «онтогенез клетки», вероятно, первым использовал Бауэр (1935). Ученик Баэра - Токин (1939) обратил внимание, что особь (томит), образованная в результате деление брюхоресничной инфузории, получает разный набор цирр (ресничных пучков) и, соответственно, должна восстановить недостающие пучки. Токин трактовал такой процесс как онтогенез, а последний этап формирования нового ресничного аппарата он считал рекапитуляциями.
Инфузории, в отличие от других протистов, имеют клеточное тело с хорошо заметными, даже на светооптическом уровне, внешними структурами. В первую очередь – это клеточный рот, для инфузории характерно поперечное деление, в результате которого одна из дочерних клеток одна из дочерних клеток получает цистом (и соответствующий ресничный аппарат), а другая должна его достраивать. Как оказалось, построение нового ротового аппарата (стоматогенез) протекает в разных группах инфузорий по – разному. Последовательность процессов, которые протекают в стоматогенезе инфузорий, трактуются как онтогенез.
Этапы стоматогенеза рассматриваются в качестве рекапитуляций.
Корлисс (Corliss, 1968) полагает, что во многих таксонах простейших можно найти примеры рекапитуляций предковых признаков. Особенно это касается групп со сложно сформированными наружными структурами: пелликулой или различного рода скелетными образованиями. По его мнению, такие примеры можно найти у миксоспоридий (скульптура стенок спор), грегарин (мембрана гаметоцист) и т.д.
И. В. Довгаль, в отличие от других протистологов считает, что стоматогенез инфузорий и ранние этапы метаморфоза расселительных стадий сидячих инфузорий (и аналогичные процессы у других групп одноклеточных) – проявление не биогенетического закона, а закона зародышевого сходства Бэра (Довгаль, 2000; Dovgal, 2002). За основу он принимает формулировку закона зародышевого сходства из монографиии И. И. Шмальгаузена (1969).
Биогенетический закон не справедлив для одноклеточных. Для протистов справедлив закон зародышевого сходства Бэра.
